Genetika - tirik organizmlarning irsiyat va o'zgaruvchanligi haqidagi fan. Klassik genetikadan farqli o'laroq, bakterial genetika mikrobiologiyaning nisbatan yosh bo'limi bo'lib, birinchi ishlar 1940-yillarning boshlarida paydo bo'lgan. Yaratilganidan beri va hozirda u tabiatshunoslikning ustuvor yo'nalishi bo'lib kelgan. Bu, birinchidan, bakterial genetikaning asosiy tamoyillari ko'p hujayrali yoki bir hujayrali bo'lishidan qat'i nazar, barcha hayvon va o'simlik organizmlariga bir xil darajada tatbiq etilishi bilan bog'liq. Ikkinchidan, irsiyat tabiatini o'rganishda prokariotlar asosan Drosophila meva pashshasi, sichqoncha va makkajo'xori kabi sevimli genetika ob'ektlarini almashtirdilar, chunki bakteriyalar yuqori ko'payish tezligiga ega va yuqori zichlikdagi populyatsiyalarni hosil qiladi va ularni etishtirish jarayoni. nisbatan oddiy. Bu genetik tahlilni muhim joyni egallamaydigan juda ko'p sonli shaxslardan foydalangan holda qisqa vaqt ichida o'tkazish imkonini beradi.
Bakterial genetikani o'rganish ham shubhasiz amaliy qiziqish uyg'otadi, masalan, patogen xususiyatlarni uzatish mexanizmlarini o'rnatish va dori vositalariga qarshilik va boshqalar. Bakteriyalar genetikasi patogenning DNK va RNKsini aniqlash asosida yuqumli kasalliklarni tashxislashning zamonaviy usullarini ishlab chiqishga imkon berdi.
Вы уже знаете о суперспособностях современного учителя?
Тратить минимум сил на подготовку и проведение уроков.
Быстро и объективно проверять знания учащихся.
Сделать изучение нового материала максимально понятным.
Избавить себя от подбора заданий и их проверки после уроков.
Просмотр содержимого документа
«Bakterialar genatikasi»
D. R. Qo’ldasheva, M. I. Nematov
O‘ZBЕKISTON RЕSPUBLIKASI OLIY VA O‘RTA MAXSUS
TA’LIM VAZIRLIGI O‘ZBЕKISTON RЕSPUBLIKASI SOG‘LIQNI SAQLASH VAZIRLIGI BUXORO TIBBIYOT INSTITUTI HUZURIDAGI ABU ALI IBN SINO NOMIDAGI JAMOAT SALOMATLIGI TEXNIKUMI
Umumkasbiyfanlar kafedrasi
BAKTERIYALAR GENETIKASI
(Texnikum o’quvchilari uchun o’quv uslubiy qo’llanma)
BUXORO-2025
Tuzuvchilar: D. R. Qo’ldasheva - Abu Ali ibn Sino nomidagi jamoat salomatligi texnikumi “Umumkasbiy fanlar” kafedrasi o’qtuvchisi.
M. I. Nematov - Abu Ali ibn Sino nomidagi jamoat salomatligi texnikumi “Umumkasbiy fanlar” kafedrasi o’qtuvchisi.
Taqrizchilar:O. I. Jabborova - Kafedra mudiri “Tibbiy biologiya kafedrasi” kafedrasi mudiri dotsent
Uslubiy qo’llanma institut markaziy uslubiy komissiyasining ____”____” 2025 yil_____- son majlis yig’ilishi bayonida ko’rib chiqildi.
MUK raisi, dotsent: Jarilkasinova G.J
Institut ilmiy kengashining ____”____” 2025 yil_____- son yig’ilish bayonida ko’rib chiqildi.
Ilmiy kotib, PhD: Do’stova N.Q
Tarkib
Kirish. 4
Fenotipik va genotipik o'zgaruvchanlik. 8
Bakteriyalardagi mutatsiyalar 9
Bakteriyalarda rekombinatsiya 11
Bakteriyalarda genetik almashinuv turlari 11
Xulosa 28
Kirish.
Genetika - tirik organizmlarning irsiyat va o'zgaruvchanligi haqidagi fan. Klassik genetikadan farqli o'laroq, bakterial genetika mikrobiologiyaning nisbatan yosh bo'limi bo'lib , birinchi ishlar 1940-yillarning boshlarida paydo bo'lgan. Yaratilganidan beri va hozirda u tabiatshunoslikning ustuvor yo'nalishi bo'lib kelgan. Bu, birinchidan, bakterial genetikaning asosiy tamoyillari ko'p hujayrali yoki bir hujayrali bo'lishidan qat'i nazar, barcha hayvon va o'simlik organizmlariga bir xil darajada tatbiq etilishi bilan bog'liq. Ikkinchidan, irsiyat tabiatini o'rganishda prokariotlar asosan Drosophila meva pashshasi, sichqoncha va makkajo'xori kabi sevimli genetika ob'ektlarini almashtirdilar, chunki bakteriyalar yuqori ko'payish tezligiga ega va yuqori zichlikdagi populyatsiyalarni hosil qiladi va ularni etishtirish jarayoni. nisbatan oddiy. Bu genetik tahlilni muhim joyni egallamaydigan juda ko'p sonli shaxslardan foydalangan holda qisqa vaqt ichida o'tkazish imkonini beradi.
Bakterial genetikani o'rganish ham shubhasiz amaliy qiziqish uyg'otadi, masalan, patogen xususiyatlarni uzatish mexanizmlarini o'rnatish va dori vositalariga qarshilik va boshqalar. Bakteriyalar genetikasi patogenning DNK va RNKsini aniqlash asosida yuqumli kasalliklarni tashxislashning zamonaviy usullarini ishlab chiqishga imkon berdi.
Bakteriyalarning genetik apparatini tashkil etish.
Prokaryotlarning genetik materiali ixcham tuzilishda joylashgan va sitoplazmaning cheklangan joylarida joylashgan DNK molekulasi (molekulalari) bilan ifodalanadi, eukaryotlardan farqli o'laroq, o'z yadro membranasiga ega emas. Ushbu xususiyatlarni hisobga olgan holda, prokaryotlarning genetik apparati odatda deyiladi nukleoid . DNK molekulasi halqa shaklida yopiq bo'lib, chiziqli joylashgan bir necha ming genlarni o'z ichiga oladi va xromosoma deb ataladi. Bakterial xromosoma bakterial hujayra uchun muhim funktsiyalarni kodlaydi.
Bakterial xromosomaning kattaligi turli bakteriyalarda farq qiladi. Misol uchun, E. coli bakterial xromosomada 4,7x106 bp mavjud. U 4300 ga yaqin genni o'z ichiga oladi. Taqqoslash uchun: viruslarning DNK hajmi taxminan 103 bp, xamirturush - 107 bp va inson xromosoma DNKsining umumiy uzunligi 3x109 bp.
Ko'pgina bakterial turlardagi har bir bakterial hujayra bitta xromosomani o'z ichiga olsa ham (aksariyat bakteriyalar haploid organizmlar), ko'pincha intensiv o'sayotgan madaniyatda hujayradagi DNK miqdori 3, 4, 8 yoki undan ko'p xromosomaga teng massaga yetishi mumkin. Bundan kelib chiqadiki, "nukleoid" va "xromosoma" atamalari har doim ham bir-biriga mos kelmaydi. Sharoitga qarab, bakteriya xujayrasining nukleoidi bir xil xromosomaning bir yoki bir necha nusxasidan iborat bo'lishi mumkin.
o'z genomida ekstraxromosoma (plazmidlar) va migratsiya ( IS ketma-ketliklari va transpozonlar) elementlarni o'z ichiga olishi mumkin .
Plazmidlar bakteriyalarning xromosomadan tashqari mobil qo'shimcha genetik tuzilmalari bo'lib, ular ikki zanjirli DNK molekulalari bo'lib, hajmi 103 dan 106 bp gacha. Ular halqa shaklida yoki chiziqli bo'lishi mumkin.
Plazmidlar avtonom nusxa ko'chirish (ko'paytirish) va hujayra sitoplazmasida mavjud bo'lishi mumkin, shuning uchun hujayrada plazmidlarning bir nechta nusxalari bo'lishi mumkin. Plazmidlar, shuningdek , xost xromosomasiga kiritilishi ( integratsiyasi ) va u bilan birga ko'payishi mumkin. Uzatish va uzatmaslik mavjudtransmissiya plazmidlari. Transmissiv (konjugativ) konjugatsiya jarayonida bir bakteriyadan ikkinchisiga o'tishga qodir. O'tkazuvchanlik faqat plazmidlarni uzatish uchun mas'ul bo'lgan genlarni birlashtirgan tra operonga ega bo'lgan yirik plazmidlarga xosdir . Bu genlar jinsiy pili kodlaydi, ular o'tkazuvchan plazmidni o'z ichiga olmaydi, hujayra bilan ko'prik hosil qiladi, ular bo'ylab plazmid DNKsi yangi hujayraga o'tkaziladi.
Tra genlarini tashmaydigan kichik plazmidlar o'z-o'zidan uzatilishi mumkin emas, lekin ularning konjugatsiya apparati yordamida transmissiv plazmidlar ishtirokida uzatishga qodir. Bunday plazmidlar mobilizatsiya, jarayonning o'zi esa o'tkazilmaydigan plazmidning mobilizatsiyasi deb ataladi.
Plazmidlar bakteriya hujayrasining hayoti uchun muhim bo'lmagan, ammo noqulay yashash sharoitlariga duchor bo'lganda bakteriyalarga afzallik beradigan funktsiyalarni kodlaydi.
Plazmidlar tomonidan bakteriya hujayrasiga beriladigan fenotipik xususiyatlar orasida quyidagilar ajralib turadi:
• antibiotiklarga qarshilik;
• patogenlik omillarini ishlab chiqarish;
• antibiotik moddalarni sintez qilish qobiliyati;
• kolitsinlarning hosil bo'lishi;
• murakkab organik moddalarning parchalanishi;
• cheklash va modifikatsiyalash fermentlarini hosil qilish.
Tibbiy mikrobiologiyada bakteriyaning antibiotiklarga chidamliligini ta'minlovchi plazmidlar alohida ahamiyatga ega bo'lib, ular R - plazmidlar (inglizcha qarshilik - qarshi ta'sir) deb ataladi va qonda yuqumli jarayonning rivojlanishiga hissa qo'shadigan patogenlik omillarini ishlab chiqarishni ta'minlaydigan plazmidlardir. makroorganizm. R -plazmidlarda antibakterial dorilarni (masalan, antibiotiklar) yo'q qiluvchi fermentlar sintezini belgilovchi genlar mavjud. Bunday plazmidning mavjudligi natijasida bakteriya xujayrasi dori vositalarining butun guruhining, ba'zan esa bir nechta dori vositalarining ta'siriga chidamli (chidamli) bo'ladi. Ko'pgina R -plazmidlar o'tkazuvchan bo'lib, bakteriya populyatsiyasi bo'ylab tarqalib, uni antibakterial dorilar ta'siriga kirishib bo'lmaydi. R -plazmidlarni tashuvchi bakterial shtammlar ko'pincha kasalxona ichidagi infektsiyalarning etiologik agentlari hisoblanadi.
Patogenlik omillarining sintezini aniqlaydigan plazmidlar hozirgi vaqtda odamning yuqumli kasalliklarini qo'zg'atuvchisi bo'lgan ko'plab bakteriyalarda topilgan. Shigelloz, yersinioz, vabo, kuydirgi, ixodid borellioz va ichak escherixiozi patogenlarining patogenligi patogen plazmidlarning mavjudligi va faoliyati bilan bog'liq.
Ba'zi bakteriya hujayralarida boshqa bakteriyalarga qarshi bakteritsid ta'sir ko'rsatadigan moddalarning sintezini aniqlaydigan plazmidlar mavjud. Misol uchun, ba'zi E. coli koliform bakteriyalarga qarshi mikrobisidal faollikka ega bo'lgan kolitsinlarning sintezini aniqlaydigan Col plazmidiga ega . Bunday plazmidlarni tashuvchi bakteriya hujayralari ekologik bo'shliqlarni kolonizatsiya qilishda afzalliklarga ega.
F plazmid yoki jinsiy omil bakteriyalarning konjugatsiya va F pili hosil qilish qobiliyatini aniqlaydi . HLY plazmidi gemolizinlar sintezini kodlaydi va shunga mos ravishda eritrotsitlarni gemolizlash qobiliyatini aniqlaydi.
Plazmidlar insonning amaliy faoliyatida, xususan, gen injeneriyasida maxsus rekombinant bakteriya shtammlarini yaratishda qo'llaniladi. Bunday plazmidlar vektorlar deyiladi .
Migratsiya qiluvchi (harakatlanuvchi) genetik elementlar bakteriya genomida ham bakterial xromosomada, ham plazmidlarda uchraydi. Mobil genetik elementlarga kiritish ketma-ketligi va transpozonlar kiradi.
Qo'shish ketma-ketligi - IS elementlari (inglizcha kiritishdan ketma-ketliklar ) DNKning bir joydan ikkinchi joyga bir butun sifatida harakatlanishi mumkin bo'lgan bo'limlari. IS elementlarining o'lchami 1000 bp. Ular faqat tnp genlarini o'z ichiga oladi, bu ularning harakati yoki transpozitsiyasi uchun zarur bo'lgan transpozaza fermenti sintezini kodlaydi. IS elementlarining uchlarida teskari takrorlanishlar (ITR) mavjud (teskari teskari degan ma'noni anglatadi, ya'ni nukleotidlarning turli uchlarida joylashishi teskari yoki qarama-qarshi yo'naltirilgan). Invertli takrorlar ham ishtirok etadi va transpozitsiya uchun muhimdir. IS elementining tuzilishini sxematik tarzda quyidagicha tasvirlash mumkin:
1-rasm. IS elementining tuzilishi .
IS elementlari genomning bir hududidan ikkinchisiga, xususan, bakterial xromosomadan plazmidga va aksincha, genomning turli hududlariga kiritilgan holda plazmiddan plazmidga o'tishi mumkin. Harakatlanayotganda ular bir gen ichida integratsiyalashishi va uni faolsizlantirishi yoki tartibga solishini o'zgartirishi mumkin.
Transpozonlar murakkab harakatlanuvchi elementlardir. Ularning IS elementlaridan farqi shundaki, ular transpozitsiya uchun mas'ul bo'lgan genlardan tashqari, transpozitsiya jarayoni bilan bog'liq bo'lmagan, ya'ni har qanday fenotipning namoyon bo'lishi uchun javobgar bo'lgan funktsiyalarni aniqlaydigan strukturaviy genlarni o'z ichiga oladi. Transpozonlar antibiotiklar va og'ir metallar ionlariga chidamliligini, laktoza, rafinozani, toluol degradatsiyasini, enterotoksin sintezini va boshqalarni katabolizatsiya qilish qobiliyatini nazorat qila oladi. Shuning uchun ularni IS elementlariga qaraganda aniqlash osonroq. Transpozonlarning uzunligi 2000 bp dan ortiq. IS elementlari singari, transpozonlarda terminal takrorlash (ITR) mavjud bo'lib, ular ko'pincha IS elementlari hisoblanadi.
Ko'chma genetik elementlarning replikon bo'ylab yoki replikonlar orasidagi harakati quyidagilarga olib keladi:
• DNKning o'sha bo'limlari genlarini, ular ko'chib o'tgandan so'ng, integratsiyalashgan holda inaktivatsiya;
• genetik materialning zararlanishini shakllantirish;
• bakterial populyatsiyada genlarning tarqalishi, bu populyatsiyaning biologik xususiyatlarining o'zgarishiga, yuqumli kasalliklar qo'zg'atuvchilarining o'zgarishiga olib kelishi mumkin, shuningdek, mikroblar orasida evolyutsiya jarayonlariga yordam beradi.
fenotip shaklida namoyon qiladi . Genomda kodlangan barcha funktsiyalar fenotipda namoyon bo'lishi mumkin emas, bu yoki boshqa genlarning ba'zilari bostirilgan ("jim" genlar).
Gen - bu irsiyatning elementar zarrasi bo'lib, u o'ziga xos polipeptid sintezini yoki mos keladigan RNK molekulasining replikatsiyasini aniqlaydigan DNKning kichik qismidir. Strukturaviy genlar mavjud - ular o'ziga xos polipeptid zanjirlari va tartibga soluvchi genlar sintezini kodlaydi , ular strukturaviy genlarning faolligini nazorat qiladi. Xuddi shunday funktsiyalarga ega bo'lgan tizimli va mos keladigan tartibga soluvchi genlar majmuasi operon deb ataladi.
Hozirgi vaqtda ko'plab bakteriyalarning genetik kodi deşifrlangan. Ular uchun genlarning joylashuvi ketma-ketligini ko'rsatadigan genetik xaritalar tuzilgan .
Fenotipik va genotipik o'zgaruvchanlik.
Bakteriyalar turining asosiy xususiyatlarini saqlaydigan irsiyat bilan bir qatorda, bakteriyalar o'zgaruvchanlik bilan ajralib turadi, ya'ni. yangi xususiyatlarga ega bo'lish va mavjud bo'lganlarning bir qismini yo'qotish qobiliyati. Bundan tashqari, ularning o'zgaruvchanlik qobiliyati yuqori organizmlarga qaraganda ancha aniq. Bu ko'payishning yuqori tezligiga, mutatsiyalar chastotasiga va genetik ma'lumotlar almashinuvining turli shakllarining mavjudligiga bog'liq.
Fenotipik va genotipik o'zgaruvchanlik mavjud.
Fenotipik o'zgaruvchanlik bilan bir yoki bir nechta belgilarda o'zgarish sodir bo'ladi va faqat genomda mavjud bo'lgan ma'lumotlar bosilgan ("jim" genlar), genotip esa ta'sirlanmagan holda qoladi. Fenotip o'zgarishlari tashqi muhit (oziq muhitning tarkibi, genomga zarar keltirmaydigan turli xil kimyoviy va fizik omillarning ta'siri) ta'sirida sodir bo'ladi. Fenotipik (modifikatsiya) o'zgaruvchanlik moslashuvchan bo'lib, bakteriyalarning yashash sharoitlari o'zgarganda ularning hayotiyligini saqlab qolish imkonini beradi . Olingan modifikatsiyalar meros bo'lib qolmaydi va uni keltirib chiqargan omilning ta'siri to'xtatilgandan so'ng tez orada yo'qoladi.
Genotipik o'zgaruvchanlik - bakteriyalarning genotipining o'zgarishi natijasida ularning xususiyatlarining doimiy o'zgarishi. O'zgaruvchanlikning bu shakli meros bo'lib, uzoq muddatli. Bu mutatsiyalar yoki genetik almashinuv tufayli yuzaga kelishi mumkin.
Bakteriyalardagi mutatsiyalar
Mutatsiyalar rekombinatsiya bilan bog'liq bo'lmagan individual DNK nukleotidlari ketma-ketligidagi irsiy o'zgarishlar bo'lib, ular fenotipik ravishda bakterial hujayra morfologiyasining o'zgarishi, o'sish omillariga, masalan, aminokislotalar, vitaminlar, ya'ni talablarning paydo bo'lishi kabi ko'rinishlarga olib keladi. auksotrofiya, antibiotiklarga chidamlilik, harorat sezuvchanligining o'zgarishi, virulentlikning pasayishi (attenuatsiya) va boshqalar.
Mutatsiyalarning har xil turlari mavjud. Kelib chiqishiga ko'ra mutatsiyalar o'z-o'zidan paydo bo'ladigan va induktsiyalanganlarga bo'linadi . Spontan mutatsiyalar bakteriya populyatsiyasida eksperimental aralashuvsiz yuzaga keladigan mutatsiyalardir. Odatda, spontan mutatsiyalar DNK replikatsiyasi paytida tasodifiy xatolar tufayli yuzaga keladi. Bunday mutatsiyalar juda kam uchraydi. O'rtacha, spontan reaktsiyalarning chastotasi 10 -4 -10 -10 ni tashkil qiladi . Induktsiyalangan mutatsiya muayyan omillarga ta'sir qilganda paydo bo'ladi - mutagen agentlar ( mutagenlar ), bu mutatsiyalar chastotasini sezilarli darajada oshiradi. Mutagenlar kimyoviy, fizik va biologik omillar bo'lishi mumkin. Bularga UV nurlari, ionlashtiruvchi nurlanish, azot kislotasi, nitrozoguanidin, azotli asoslarning analoglari, ba'zi antibiotiklar, akridin bo'yoqlari, oltingugurt xantallari, transpozonlar, kiritish elementlari va boshqalar kiradi.
Barcha mutagenlar bakterial DNKga ta'sir qiladi, ammo ularning harakati o'ziga xos emas, ya'ni. Har qanday mutagen yordamida faqat ma'lum turdagi mutatsiyani ajratib olish mumkin emas. Mutagenlar faqat mutatsiyalar chastotasini oshirishi mumkin.
Fenotipik oqibatlarga ko'ra, ular bo'linadi to'g'ridan-to'g'ri va teskari . Hujayra funktsiyasining yo'qolishiga yoki o'zgarishiga olib keladigan mutatsiyaga to'g'ridan-to'g'ri mutatsiya deyiladi. To'g'ridan-to'g'ri mutatsiyalar yovvoyi turdan farq qiladigan g'ayritabiiy fenotipni keltirib chiqaradi. Shunday qilib, bular yovvoyi turdan mutantga mutatsiyalardir. Masalan , E. Coli laktoza fermentatsiyasiga qodir ( lak + fenotip) va shuning uchun lak + → lak mutatsiyasi to'g'ridan - to'g'ri bo'ladi. Olingan shaxslar mutantlardir . Mutantlarda asl xususiyatlarning tiklanishi mumkin - teskari (inglizcha teskari - teskari). lak - → lak + - teskari mutatsiya . Teskari mutatsiyalar natijasida paydo bo'lgan shaxslar revertantlar deb ataladi .
haqiqiy yoki ikkilamchi bo'lishi mumkin . Haqiqiy mutatsiyalar natijasida asl genotip tiklanadi, ya'ni. Birinchi mutatsiyada o'zgargan nukleotidlarning uchligi yana avvalgidek bir xil aminokislotalarni kodlaydi. Biroq, birinchi mutatsiyaning ta'siri genning boshqa qismida qoplanishi mumkin. Bunday mutatsiyalar deyiladi ikkilamchi . Ba'zida mutatsiyalar boshqa qo'shni genlarda ham yuzaga keladi, bu birinchi mutatsiyaning ta'sirini qandaydir tarzda chetlab o'tishga yordam beradi. Ular supressor deb ataladi . Supressor mutatsiyalar mutantning asl holatini tiklamasdan, faqat mutantning fenotipini tiklaydi.
Fenotipik ko'rinishlarga ko'ra (o'zgargan arvohning tabiati) mutatsiyalar quyidagilarga bo'linadi:
morfologik mutatsiyalar - ba'zi morfologik belgilarning o'zgarishi (masalan, kapsulali → kapsulali bo'lmagan va boshqalar);
biokimyoviy mutatsiyalar:
-auksotrof mutantlar (qo'shimcha o'sish omillarini talab qiladi);
-katabolik fermentlarning konstitutsiyaviy sintezi bo'lgan mutantlar (buzilgan tartibga solish);
- anabolik fermentlarning konstitutsiyaviy sinteziga ega mutantlar;
- virulent mutantlar;
- antigenik xossalari o'zgargan mutantlar.
Genetik strukturaning o'zgarish darajasiga ko'ra, mutatsiyalarning quyidagi turlari ajratiladi: nuqta mutatsiyalari, zarar bir juft nukleotidlar bilan chegaralanganda va xromosoma mutatsiyalari - alohida DNK bo'laklarida katta o'zgarishlar. Xromosoma mutatsiyalari quyidagi turlarda bo'lishi mumkin: deletsiya, duplikatsiya, inversiya va translokatsiya.
Deletsiya DNKning bir qismini (bir nechta nukleotidlarni) yo'qotishdir. Bunday mutatsiyalar qaytarilmas va revertantlarni olish mumkin emas. Duplikatsiya bilan xromosomaning bir qismi takrorlanadi, dislokatsiya bilan bir nechta asoslar ketma-ketligi o'zgaradi va inversiya bilan xromosomaning bir qismi 180 gradusga aylanadi .
Nuqta mutatsiyasi - bu faqat bitta tayanch juftlikka ta'sir qiladigan mutatsiya. Bunday holda, bir juft boshqasi bilan almashtiriladi. Nuqta mutatsiyalari reversiyalarning yuqori chastotasi bilan tavsiflanadi. Baza o'rnini bosuvchi mutatsiyalar ko'pincha noto'g'ri mutatsiyalar (ma'no o'zgarishi bilan mutatsiya ) bo'lib chiqadi , bunda almashtirishdan keyin bazalarning kodlash uchligi ketma-ketligi boshqa aminokislotalarni kodlaydi. Asosiy almashtirish mutatsiyalarining muhim qismi bema'ni mutatsiyalar bo'lib , aminokislotalarni kodlaydigan triplet hech qanday aminokislotalarni kodlamaydigan tripletga aylanishi bilan tavsiflanadi.
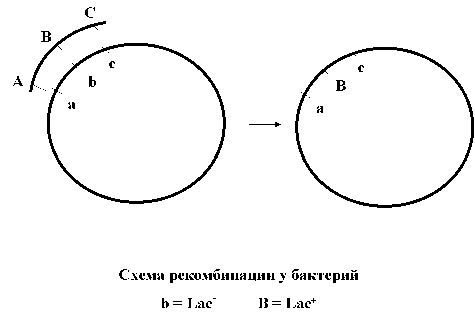
Bakteriyalarda rekombinatsiya
2-rasm. Bakteriyalarda rekombinatsiya sxemasi
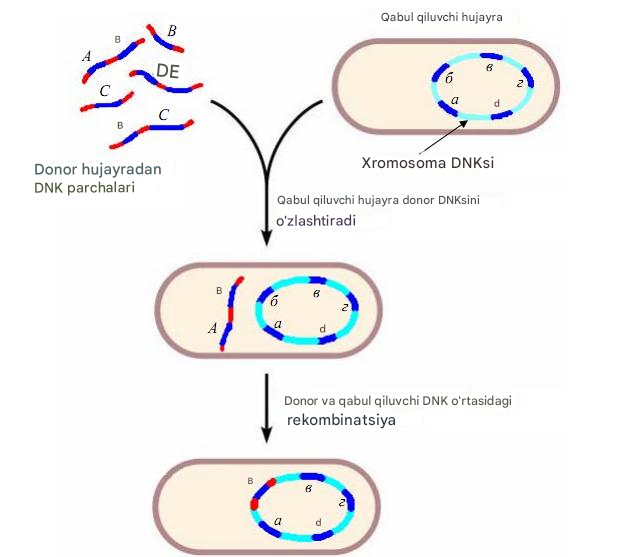
Genetik rekombinatsiya - bu donor bakteriya va retsipient bakteriyadan xossalarga ega bo'lgan yangi rekombinant DNK molekulasining shakllanishiga olib keladigan turli xil genotiplarning ikkita DNK molekulasi o'rtasidagi o'zaro ta'sir jarayoni. Bakteriyalardagi rekombinatsiya jarayoni ularning genetik apparati xususiyatlari va genetik almashinuv shakllari bilan bog'liq ba'zi farqlarga ega. Genetik transfer jarayoni qabul qiluvchi bakteriya va donor bakteriyani o'z ichiga oladi. Ularning ishtirok etish darajasi notekis, faqat donor bakteriyaning ekzogen DNK qismi qabul qiluvchi hujayraga kiradi. Bakteriyalarda rekombinatsiyaning barcha bosqichlari fermentlar tomonidan ta'minlanadi: cheklovchi fermentlar, ligazalar va boshqalar. Bakteriyalarda rekombinatsiyaning uch turi ajralib turadi: umumiy ("qonuniy"), "noqonuniy" va "saytga xos " . Umumiy yoki gomologik rekombinatsiya, agar o'zaro ta'sir qiluvchi DNK tuzilishida gomologik hududlar mavjud bo'lsa, sodir bo'ladi. Bunday rekombinatsiya faqat yaqin turlar orasida mumkin. Uni amalga oshirish uchun "noqonuniy" rekombinatsiya o'zaro ta'sir qiluvchi tuzilmalarning muhim DNK homologiyasini talab qilmaydi. "Noqonuniy" rekombinatsiya xromosomaga tez integratsiyani ta'minlaydigan IS ketma-ketliklari va transpozonlarni o'z ichiga oladi. Shu sababli, "noqonuniy" rekombinatsiya bir xil turdagi bakteriyalar o'rtasida ham, turli tur va avlod bakteriyalari o'rtasida ham sodir bo'lishi mumkin.
Rekombinatsiyaning uchinchi turi - bu aniq belgilangan DNK ketma-ketliklari va maxsus fermentlarni talab qiladigan "saytga xos ".
Bakteriyalarda genetik almashinuv turlari
DNK o'tkazishning eng ko'p o'rganilgan uchta turi bir-biridan uni tashish usuli bilan farq qiladi: transformatsiya, transduksiya, konjugatsiya.
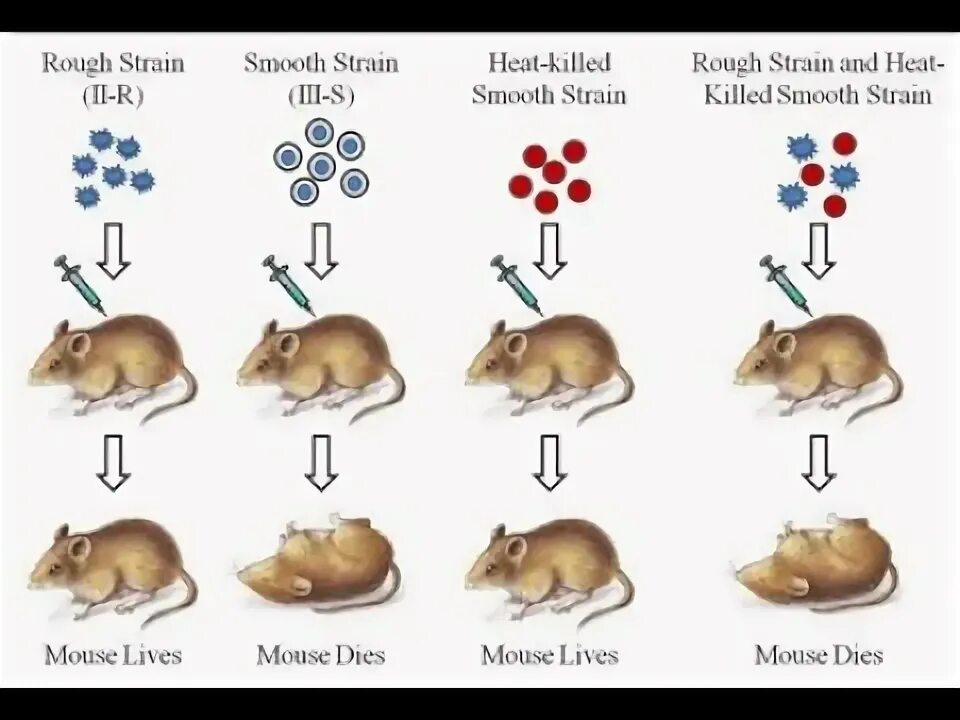
Transformatsiya - (lotincha transformatio - transformatsiya) - erkin eriydigan shaklda bakteriyalardan ajratilgan ekzogen DNK retsipient bakteriyaga o'tadi. Transformatsiya jarayonida bakteriyalarning DNKsi bir-biri bilan bog'liq bo'lsa, rekombinatsiya sodir bo'ladi. Bunday holda, o'z va tashqi kirib borgan DNKning gomologik hududlari almashinuvi mumkin. Transformatsiya birinchi marta 1928 yilda F. Griffits tomonidan qayta ishlab chiqarilgan. U bir vaqtning o'zida pnevmokokkning avirulent akapsulyar shtammlarini kiritgan va bu bakteriyalarning o'ldirilgan versiyalarini oq sichqonlarning qorin bo'shlig'iga kiritgan, buning natijasida avirulent shtammlar virulentlikka ega bo'lgan.
S - SHTAMM R - SHTAMM O’dirilgan shtamm S - va O’ldirilgan shtamm
Transformatsiya jarayoni tabiatda o'z-o'zidan sodir bo'lishi mumkin bakteriyalarning ayrim turlari , B. subtilis , H gripp , S pneumoniae , o'lik hujayralardan ajratilgan DNK qabul qiluvchi hujayralar tomonidan qabul qilinganda. Induktsiyalangan transformatsiya genetik xususiyatlari sinov madaniyatiga o'tkazilishi kerak bo'lgan bakteriyalar madaniyatidan olingan tozalangan DNK bakterial madaniyatga qo'shilganda sodir bo'ladi.
Muvaffaqiyatli transformatsiyani amalga oshirish uchun quyidagi shartlarga rioya qilish kerak:
Donorning DNKsi retsipient (yoki yaqin qarindosh) bilan bir xil turdagi bakterial madaniyatdan ajratilishi kerak.
Transformatsiya qiluvchi DNK hududi ikki torli o'ta sarmalni saqlab turishi kerak. Buning sababi shundaki, DNKning faqat bitta zanjiri retsipient hujayra ichiga kirib boradi, ikkinchisi - hujayra membranasida - qolgan zanjirning hujayra ichiga kirib borishi uchun zarur bo'lgan energiya ajralib chiqishi bilan degradatsiyaga uchraydi.
DNK konsentratsiyasi har ikki holatda ham past yoki ortiqcha bo'lmasligi kerak, rekombinantlar soni kamayadi; Transformatsiya qiluvchi DNKning yuqori molekulyar og'irligi rekombinatsiya ehtimolini oshiradi, chunki hujayra ichida transformatsiya qiluvchi DNK zanjiri endonukleazlarga ta'sir qiladi.
Qabul qiluvchi hujayralar malakali bo'lishi kerak. Kompetentlik - bakterial hujayraning DNKni singdirish qobiliyati. Bu hujayra membranasida DNKga o'ziga xos yaqinlikka ega bo'lgan maxsus oqsillarning mavjudligiga bog'liq. Gram-musbat bakteriyalarda kompetentsiya holati o'sish egri chizig'ining ma'lum fazalari bilan bog'liq (eng katta kompetentlik o'sishning logarifmik bosqichida kuzatiladi). Gram-manfiy bakteriyalarda malaka holati bakteriyalarni termal yoki elektr toki urishi bilan sun'iy ravishda yaratish kerak.
Transformatsiya to'rt bosqichda sodir bo'ladi.
Qabul qiluvchi hujayrada donor DNKsining adsorbsiyasi. DNK faqat vakolatli hujayralar kirishi mumkin bo'lgan sirt joylariga bog'lanadi.
Qabul qiluvchi hujayraga DNK fragmentlarining kirib borishi. Bu jarayon DNK zanjirlaridan birini yo'q qilish va keyingi kirish uchun zarur bo'lgan energiyani chiqarish bilan birga keladi.
DNKning qabul qiluvchi xromosomaning gomologik mintaqasi bilan bog'lanishi, so'ngra rekombinatsiya
4-rasm. Bakteriyalarda transformatsiya sxemasi.
Shuni ta'kidlash kerakki, bakteriyalarning ba'zi turlari, masalan, pnevmokokklar, har xil turdagi DNKni va hatto yuqori organizmlarning DNKsini adsorbsiyalash va o'zlashtirishga qodir.
Transformatsiyani qo'llash:
- bakterial xromosomani transformatsiya chastotasi bo'yicha xaritalash uchun;
- mikroorganizmlarning foydali shtammlarini loyihalash uchun;
- transformatsiya yordamida siz bakterial genomga ma'lum markerlarni kiritishingiz va kiruvchi mutatsiyalarni yo'q qilishingiz mumkin;
- transformatsiya izolyatsiya qilingan DNKda turli xil genetik va molekulyar biologik tajribalar uchun namuna bo'lib xizmat qilishi mumkin, chunki transformatsion faollik darajasi uning strukturaviy yaxlitligining juda sezgir ko'rsatkichidir. Shunday qilib, turli xil agentlarning inaktivatsiya yoki mutagen ta'sirining mexanizmlarini va ular keltiradigan zararning xarakterini o'rganish mumkin.
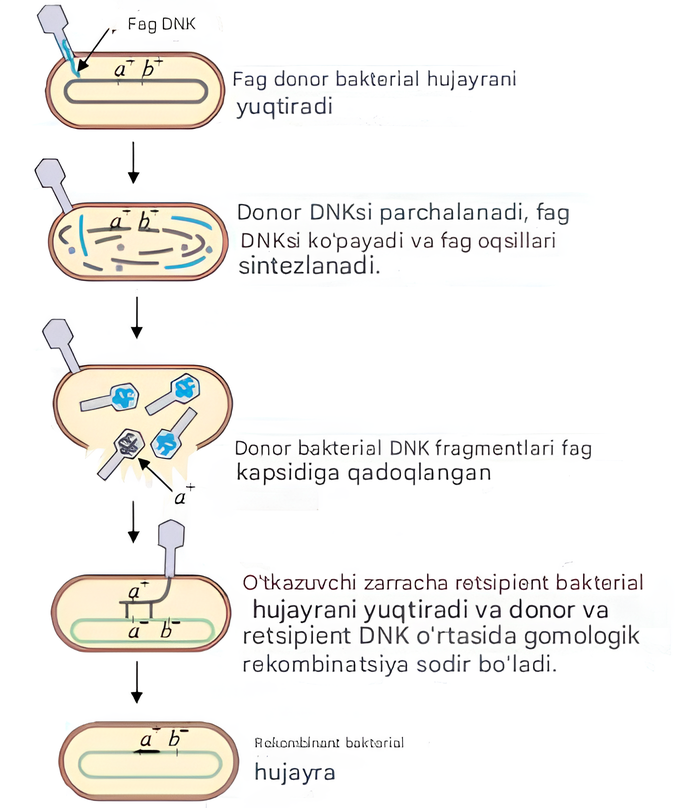
Transduktsiya- bakteriofag yordamida genetik materialni donor bakteriyadan retsipient bakteriyaga o'tkazish jarayoni. Bu jarayon 1951 yilda N. Zinder va J. Lederberg tomonidan kashf etilgan. Transduksiya vaqtida xromosoma genlari yoki plazmidlar bakteriofag boshiga qadoqlanishi kerak; fag zarrachasining bir qismi sifatida lizislangan hujayradan (donor) chiqib, yangi infektsiya akti paytida boshqa hujayraga (qabul qiluvchiga) kiradi.
Transduksiyaning uch turi mavjud: nospetsifik (umumiy), o'ziga xos va abortiv.
Umumiy (nospesifik) transduksiya - bakteriofag tomonidan bakterial xromosomaning istalgan qismining bir qismining ko'chirilishi. Bakteriofag bilan kasallangan hujayrada qiz populyatsiyasini yig'ish paytida bakterial DNK yoki plazmidning bir qismi virusli DNK bilan birga yoki uning o'rniga ba'zi faglarning boshlariga kirib borishi mumkin. Bu jarayon fag infektsiyasidan keyin bakterial DNK bo'laklari va fag DNKsi bilan bir xil o'lchamdagi bakterial DNK bo'lagi virus zarrachasiga taxminan 1000 fag zarrachasidan 1 nisbatda kirib borishi sababli yuzaga keladi. Transduktsiyaning ushbu shakli bilan deyarli har qanday genlar qabul qiluvchi hujayralarga kiritilishi mumkin. Nonspesifik transduksiya fenomeni bakterial xromosomani xaritalash uchun ishlatilishi mumkin.
5-rasm. Nospesifik transduksiya sxemasi.
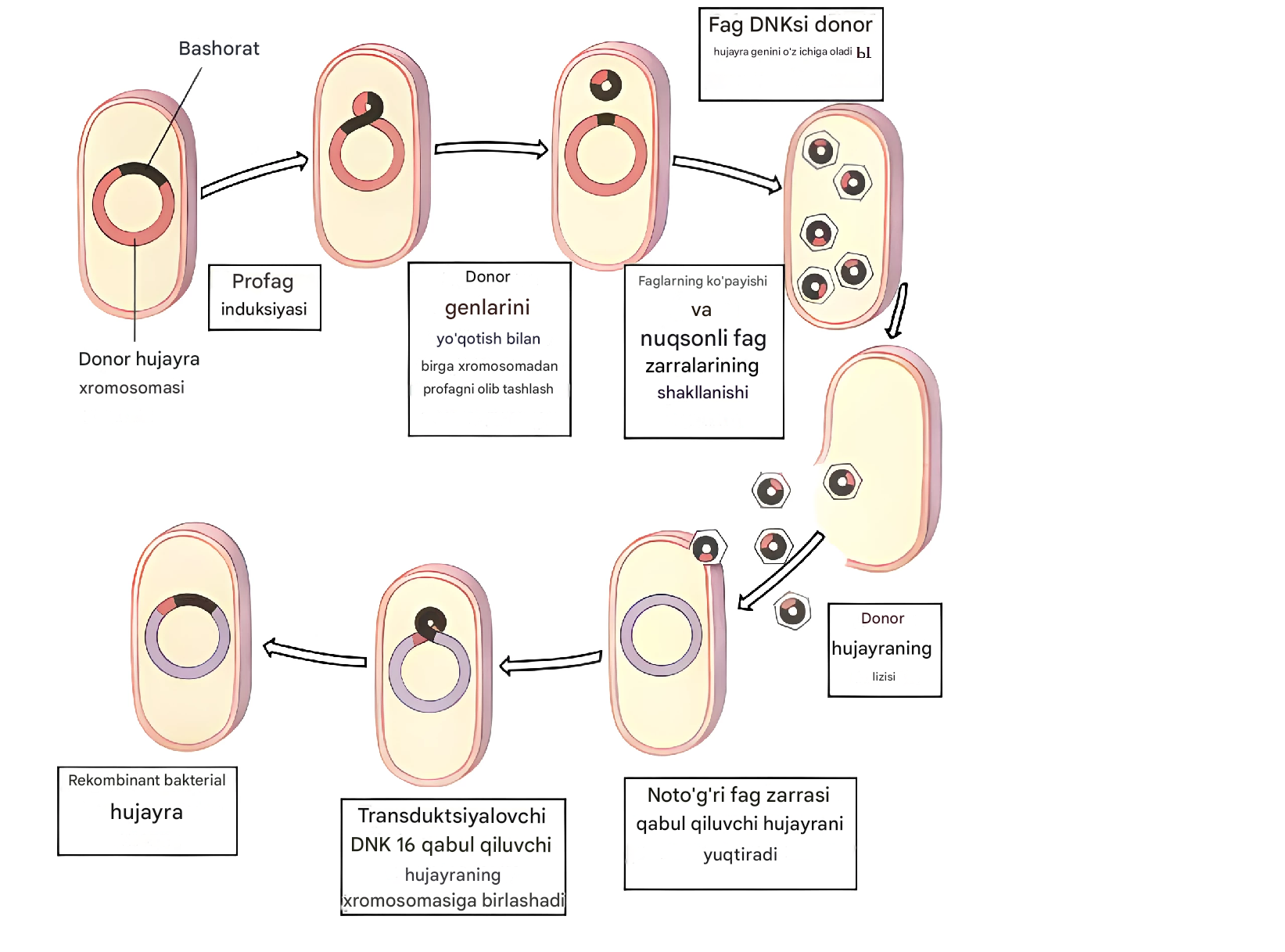
Maxsus transduksiya fag DNKsi bakteriyaga integratsiyalashganda profag hosil qilganda sodir bo'ladi. Fag DNKsi bakterial xromosomadan chiqarilganda, tasodifiy jarayon natijasida fag DNKsi qo'shilgan joyga qo'shni bakterial xromosomaning bir qismi ushlanadi. Ko'pgina mo''tadil faglar bakterial DNKga ma'lum hududlarda integratsiyalashganligi sababli, bunday bakteriofaglar donor bakterial DNKning ma'lum bir qismini qabul qiluvchi hujayraga o'tkazish bilan tavsiflanadi. Spesifik transduktsiya bakteriyalar o'rtasida virulentlik genlarini uzatish mexanizmi bo'lib xizmat qilishi mumkin, agar bu genlar profag integratsiyasi joylariga yaqin joyda joylashgan bo'lsa.
Eng tipik misol fag l tomonidan amalga oshiriladigan transduktsiyadir. U odatda ma'lum genlarni o'zgartiradi: gal (galaktoza sintezini kodlaydi) va bio (biotin sintezini kodlaydi). Profag holatiga o'tgandan so'ng, fag l mezbon bakteriya xromosomasining ma'lum bir hududiga - gal va biogenlar orasiga kiradi. Bakteriya xromosomasidan fag DNKsining ajralishi noto'g'ri sodir bo'lishi mumkin va uning bir qismi xromosomada qoladi va yaqin joylashgan genlar fag DNK tomonidan ushlanadi. Agar transduktsiyalovchi fag ma'lum bir genda nuqsonli bo'lgan hujayralarni, masalan, gal - ni yuqtirsa, rekombinatsiya rekombinant (transduktant) gal + hosil qilish uchun bakteriyalarning o'z nuqsonli genini buzilmagan transduktsiyalangan gen bilan almashtirish bilan sodir bo'lishi mumkin.
6-rasm. Maxsus transduktsiya sxemasi.
Abortiv transduktsiya . Abortiv transduksiya vaqtida kiritilgan donor DNK fragmenti retsipient xromosomasiga birlashtirilmaydi, balki sitoplazmada qoladi va u erda mustaqil ravishda ishlaydi. Keyinchalik, u qiz hujayralaridan biriga o'tadi (ya'ni, unilineally meros qilib olinadi) va keyin avlodda yo'qoladi.
Tabiiy sharoitda transduktsiya juda tez-tez sodir bo'ladi , turli bakteriofaglar bilan kasallangan bakteriyalarning keng tarqalishi tufayli. Bundan tashqari, nafaqat turlar, balki turlararo va avlodlararo transduksiya ham mavjud.
Transduktsiya kichik miqdordagi genetik materialni, odatda bir yoki ikkita yaqin bog'liq genlarni uzatadi. Transduksiya yordamida bakterial genlar, plazmidlar va transpozonlar uzatiladi, bu genetik muhandislikda qo'llaniladi.
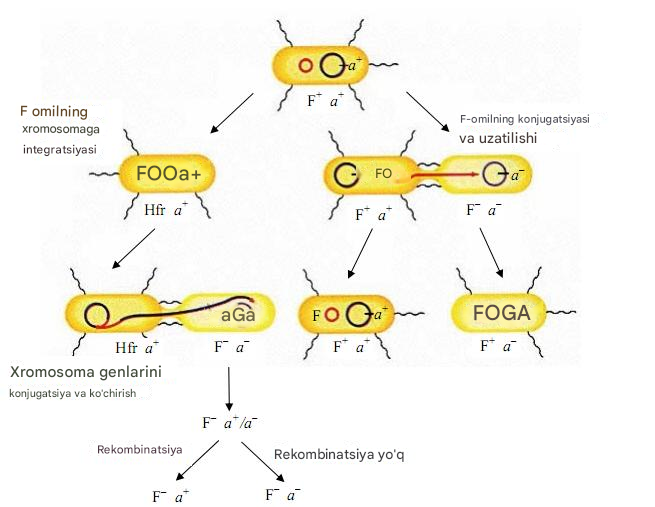
Konjugatsiya - ikki hujayra o'rtasida to'g'ridan-to'g'ri aloqa qilish orqali genetik materialning uzatilishi. Konjugatsiya birinchi marta 1946 yilda J. Lederberg va E. Tatum tomonidan kashf etilgan. Konjugatsiyaning zaruriy sharti donor hujayrada transmissiv plazmidning mavjudligidir. O'tkazuvchan plazmidlar jinsiy pili kodlaydi, ular donor hujayra va retsipient hujayra o'rtasida konjugatsiya ko'prigi hosil qiladi, bu orqali plazmid DNKsi donor hujayradan retsipient hujayraga o'tadi. Ushbu transfer natijasida qabul qiluvchi hujayra donorlik xususiyatlarini oladi. Integral transmissiv plazmid F omil hisoblanadi. F omilga ega bo'lgan donor hujayralar F + hujayralar deb ataladi va F omil bo'lmagan qabul qiluvchi hujayralar F - hujayralar deb ataladi.
7 -rasm. Bakterial konjugatsiyaning mikrografisi.
Donor hujayralarning bir necha turlari mavjud:
F plazmidlarini o'z ichiga olgan donor hujayralar F + hujayralari deb ataladi .
Xromosomaga F faktori o'rnatilgan donor hujayralar Hfr hujayralari deb ataladi (inglizcha yuqori chastota ning rekombinatsiya - rekombinatsiyaning yuqori chastotasi). Agar F omil donor hujayra xromosomasiga birlashgan bo'lsa va xromosoma bilan bitta transmissiv replikon vazifasini bajara boshlasa, u holda donor xromosomasi retsipient hujayraga o'tish qobiliyatiga ega bo'ladi.
Integratsiyalashgan holatdan F -omilli plazmid sitoplazmaga (avtonom holatga) qaytishi mumkin. Bunday holda, ba'zi xromosoma genlari plazmidga o'tkazilishi mumkin. Bunday donor hujayralar F' hujayralari deb ataladi .
8-rasm. Bakteriyalarda konjugatsiya sxemasi.
Donorning turiga qarab, o'tishning quyidagi turlari ajratiladi:
F + kesib o'tish xF-: faqat F -plazmid ko'chiriladi, F - hujayra esa F + - hujayra bo'lib , plazmid va donorning xususiyatlarini oladi. Xromosoma genlari uzatilmaydi.
Ketish HfrxF -:(rekombinantlar mavjud) bakterial genlar uzatiladi. Butun xromosoma zanjirining kirib borishi uchun ko'p vaqt kerak bo'ladi va, qoida tariqasida, to'liq o'tish kamdan-kam hollarda amalga oshiriladi, shuning uchun xromosomaning ushbu qismida joylashgan, qabul qiluvchi hujayraga kirishga ulgurmagan genlar; umuman uzatiladi. Shuning uchun, bunday kesishish paytida qabul qiluvchi hujayralar, qoida tariqasida, donorga aylanmaydi
F'x ni kesib o'tish F -:(rekombinantlar mavjud) F + kesishishiga o'xshash tarzda sodir bo'ladi x F-qabul qiluvchi hujayra esa donor hujayraga aylanadi. Ammo bu tipning o'ziga xosligi shundaki, F '-plazmid o'z genomining bir qismi sifatida bakterial xromosomaning bir qismini o'z ichiga oladi.
Konjugatsiya 4 bosqichda sodir bo'ladi.
Jinsiy villi yordamida donor hujayraning qabul qiluvchi hujayraga biriktirilishi .
Ikkala hujayra o'rtasida konjugatsiya ko'prigining shakllanishi .
DNK zanjirlaridan birini sindirish va despiratsiya qilish, konjugatsiya ko'prigi orqali proksimal uchining qabul qiluvchi hujayraga kirib borishi .
Qabul qiluvchi hujayradagi DNKning ikkinchi zanjirini to'ldirish va donor DNKni tiklash.
Konjugatsiyaning biologik ahamiyati bakteriyalarning antibiotiklarga chidamliligining tarqalishi bilan aniq ko'rsatilgan. Har 10 6 hujayra bo'linishida bir marta sodir bo'ladigan mutatsiya natijasida bakteriya antibiotiklarga chidamli bo'lishi mumkin . Biroq, bir marta o'zgartirilgandan so'ng, genetik ma'lumotlar konjugatsiya orqali o'xshash bakteriyalar orasida tez tarqalishi mumkin, chunki har uch yaqin bakteriyadan biri bu turdagi genetik transferga qodir.
6. Yuqumli kasalliklar diagnostikasining molekulyar genetik usullari.
molekulyar genetik usullarini o'rganilayotgan materialda patogenning DNK yoki RNKsini aniqlash imkonini beruvchi usullarni tushunish kerak.
Ushbu usullar molekulyar gibridizatsiya va polimeraza zanjiri reaktsiyasini (PCR, PCR ) o'z ichiga oladi.
Molekulyar gibridlanish
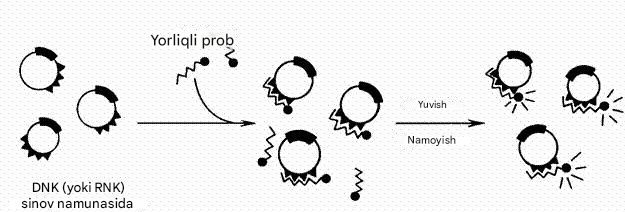
Molekulyar duragaylash usuli DNK va RNK ning sun'iy ravishda sintezlangan va ferment, ftorxrom yoki izotop bilan belgilangan komplementar oligonukleotid bo'laklari bilan maxsus birlasha olish (gibridlanish) qobiliyatiga asoslangan. Ushbu qismlarga zondlar deyiladi.
Molekulyar gibridizatsiyani amalga oshirish uchun o'rganilayotgan DNK molekulasi denatüratsiya qilinadi, bitta ip maxsus filtrga o'rnatiladi, u etiketli probni o'z ichiga olgan eritma ichiga joylashtiriladi. Ikki tomonlama spirallarning shakllanishi uchun qulay sharoitlar yaratiladi. Agar prob va tekshirilayotgan DNK o'rtasida bir-birini to'ldiruvchi bo'lsa, ular o'zaro juft spiral hosil qiladi. Gibridizatsiya tugallangandan va bog'lanmagan mahsulotlar yuvilgandan so'ng, hosil bo'lgan kompleks tegishli yorliq yordamida aniqlanadi.
9-rasm. Muayyan yorliqli zond bilan patogen DNK yoki RNKni aniqlash uchun molekulyar gibridlanish reaktsiyasining sxemasi.
Polimeraza zanjiri reaktsiyasi.
Polimeraza zanjiri reaktsiyasi (PCR) usuli 1985 yilda amerikalik tadqiqotchi Kari Mullis tomonidan taklif qilingan. Ushbu ishi uchun u 1993 yilda kimyo bo'yicha Nobel mukofotiga sazovor bo'lgan. Perinatal amaliyotda birinchi marta o'roqsimon hujayrali anemiyaning prenatal diagnostikasi uchun PCR usuli qo'llanildi. Biroz vaqt o'tgach, bu usul birinchi navbatda OIV-1ni aniqlash, keyin esa virusli va bakterial infektsiyalarning boshqa patogenlarini, shuningdek protozoa (toksoplazmoz) keltirib chiqaradigan infektsiyalarni aniqlash uchun qo'llanila boshlandi.
PCR DNKning denaturatsiya va renaturatsiya qilish qobiliyatiga va DNK zanjirlarining komplementarligiga asoslanadi.
Usulning printsipi DNK polimeraza fermenti yordamida primerlar bilan chegaralangan DNK mintaqasini ko'paytirish (kuchaytirish).
Har bir keyingi kuchaytirish sikli uchun DNKning asl qismi ham, yangi sintez qilingan fragmentlar ham (kuchaytirgichlar) ikki barobar ortadi.
Natijada, fragmentlar soni eksponensial ravishda oshadi (zanjir reaktsiyasi). 30-40 tsikldan keyin ularning soni bir necha milliarddan oshadi, bu ularni turli usullar yordamida aniqlash imkonini beradi.
Eng oddiy holatda PCRni o'tkazish uchun quyidagi komponentlar talab qilinadi:
Tekshirilgan biologik materialda yuqumli agentning aniqlanishi mumkin bo'lgan DNKsi (RNK) .
primerlar (oligonukleotidlar) 20-30 nukleotiddan iborat bo'lgan qisqa DNK zanjirlari bo'lib, DNKning ikkita zanjirining har birining 3' uchini to'ldiruvchi nukleotidlar ketma-ketligi aniqlanadi. Primerlar kiradi vitro kimyoviy sintez yoki turli patogen mikroorganizmlarning nuklein kislotalaridan ajratilgan.
Termostabil DNK polimeraza - bu DNK polimerizatsiya reaktsiyasini katalizlovchi ferment (komplementar zanjirlarning tugallanishi). PCRda foydalanish uchun polimeraza uzoq vaqt davomida yuqori haroratlarda faol qolishi kerak, shuning uchun termofillardan ajratilgan fermentlar ishlatiladi - Thermus aquaticus (Taq polimeraza), Pyrococcus furiosus (Pfu polimeraza), Pyrococcus woesei (Pwo polimeraza) va boshqalar.
Polimeraza ishlashi uchun zarur bo'lgan Mg2+ ionlari.
Kerakli reaksiya sharoitlarini ta'minlaydigan tampon eritmasi - pH , eritmaning ion kuchi .
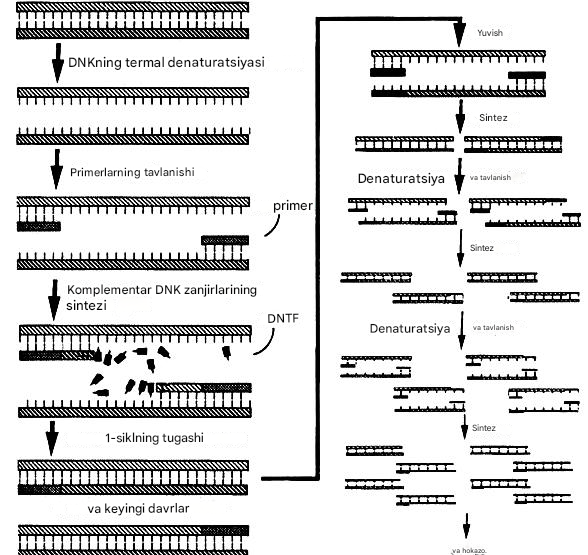
PCR tadqiqotining bosqichlari:
1. Nuklein kislotalarning izolyatsiyasi. Birinchi bosqichda barcha DNK (DNK o'z ichiga olgan mikroorganizmlar uchun) yoki RNK ( NASBA usuli uchun) ajratiladi. yoki viruslarni o'z ichiga olgan RNK) sinov materialidan.
2. PCRning o'zi yoki kuchaytirilishi. Birinchi bosqichda ajratilgan DNK nukleotidlar, PCR buferi, polimeraza va primerlar aralashmasini o'z ichiga olgan eritmaga qo'shiladi. Amplifikatsiya uch bosqichda amalga oshiriladi: denatürasyon, tavlanish, cho'zish. Birinchidan, reaktsiya aralashmasi 90-94 ° C ga qadar isitiladi va shu bilan denaturatsiyaga olib keladi - ikki zanjirli DNK zanjirlarining echilishi. Keyin harorat 50-70 ° C ga tushiriladi va primerlarni qo'shish (tavlash) boshlanadi . Primerlar tanlangan maqsadni cheklaydigan qisqa hududlarga maxsus biriktirilgan (qo'shimcha juftlik printsipiga ko'ra). Primerlar faqat ma'lum bir patogenga xos bo'lgan tanlangan DNK fragmentlariga biriktiriladi va boshqa DNK ketma-ketligi bilan o'zaro ta'sir qilmaydi (va klinik namunada ham inson, ham turli mikroorganizmlar DNKsi juda ko'p). Bu diagnostik PCR test tizimining mutlaq o'ziga xosligini ta'minlaydi. Ya'ni, dastlabki PCR eritmasidagi primerlar 100% ehtimollik bilan biriktirish uchun maqsad topadi. Primerlarning qo'shilishi natijasida "urug'" komplekslari bo'lgan tuzilmalar [DNK shabloni + primer] hosil bo'ladi. Shundan so'ng, 3' uchidan boshlab, shablonlarda bir ipli DNK bo'laklarini to'ldiruvchi sintez orqali cho'zilish bosqichi (primerlarning qurilishini yakunlash) keladi . Bu jarayon maxsus ferment - Taq polimeraza (issiqlikka chidamli DNK polimeraza) va nukleotidlar (qurilish materiali sifatida) yordamida amalga oshiriladi.
Yangi sintez qilingan ikki zanjirli DNK ham denaturatsiyaga uchraydi va aralashmada ortiqcha bo'lgan primerlarni yana bog'laydi. Olingan tuzilmalar, tugallangandan so'ng, primer ketma-ketligi bilan cheklangan o'ziga xos DNK bo'laklarini hosil qiladi. Shunday qilib, bu tsikllar takrorlanganda, primer qo'nish joylari o'rtasida joylashgan DNK bo'limining nusxalari soni eksponent ravishda ortadi.
3.Natijalarni hisobga olish. To'plangan amplifikatsiya mahsulotlari (primer qo'nish joylari orasidagi ko'p sonli DNK nusxalari) jel elektroforezi bilan aniqlanishi mumkin.
* An'anaviy PCRdan farqli o'laroq, NASBA (nuklein kislotalar ketma-ketligiga asoslangan kuchaytirish) maqsadlari mikroorganizmlar ribosomalarining RNK molekulalari bo'lib, bir qator afzalliklarni ta'minlaydi.
Rasm. 10. Qizdirish bosqichlari
PCR ning afzalliklari
1. Ko'p qirralilik. PCR har qanday biologik namunada DNKni aniqlash uchun ishlatilishi mumkin. Bundan tashqari, bu mikroorganizmlar DNKsi va inson DNKsi uchun bir xil darajada amal qiladi.
2. Yuqori o'ziga xoslik. O'ziga xoslik PCR faqat ma'lum bir patogenga xos bo'lgan genning noyob hududini aniqlashi bilan aniqlanadi. Tanlangan primerlarga qarab, PCR test tizimi bir xil turdagi mikroorganizmlarning avlodlari, turlari, serotiplari va hatto patogen va patogen bo'lmagan shtammlarini farqlashi mumkin.
3. Yuqori sezuvchanlik. Polimeraza zanjiri reaktsiyasi DNKning bitta nusxasini aniqlay oladi. O'rtacha ko'pgina zamonaviy test tizimlarining sezgirlik chegarasi DNKning 10 dan 100 nusxasini tashkil qiladi. Bu madaniy tadqiqot usullarining sezgirligidan sezilarli darajada oshadi.
4. Biologik materialning kichik hajmi. Tahlil eng kichik namuna hajmida (bir necha mikrolitrgacha) mumkin, bu pediatriya, neonatologiya, nevrologiya va sud tibbiyotida juda muhimdir.
5. Faqat o'tkir emas, balki yashirin infektsiyalarni ham tashxislash imkoniyati. PCR usuli, ayniqsa, latent va surunkali infektsiyalarda tez-tez uchraydigan mikroorganizmlarning etishtirish qiyin bo'lgan, etishtirilmaydigan va turg'un shakllarini tashxislashda samaralidir.
6.Kasalliklarni erta tashxislash qobiliyati - tadqiqot o'tkazish uchun maksimal vaqt 48 soat.
Haqiqiy vaqtda PCR
Haqiqiy vaqtda PCR (real-time PCR, monitoring PCR, RT-PCR) - maqsadli DNKni kuchaytirish bosqichlari va olingan mahsulotlarni aniqlash bir vaqtning o'zida bitta probirkada sodir bo'ladigan usul. RT-PCR o'rganilayotgan materialdagi DNK tarkibini miqdoriy baholash imkonini beradi. Ushbu usulni o'rnatish uchun namunadan chiqadigan lyuminestsent nurlanishni ro'yxatga olish uchun o'rnatilgan optik blokli maxsus kuchaytirgich talab qilinadi. Shuningdek, PCR kuchaytirgichining ishlashini qo'llab-quvvatlaydigan va polimerizatsiya reaktsiyasi va yangi sintezlangan DNK ketma-ketliklarining to'planishini tavsiflovchi grafikni tuzadigan kompyuter dasturlari. Eng oddiy shaklda, PCR mahsulotlarining to'planishi er-xotin zanjirli DNK bilan bog'langan floresan bo'yoqlar yordamida aniqlanadi. Shunday bo'yoqlardan biri SYBR Green hisoblanadi. Erkin holatda, bu bo'yoq tomonidan hosil bo'lgan floresans nisbatan past bo'ladi, lekin bo'yoq ikki ipli DNK bilan bog'langanda, floresans sezilarli darajada kuchayadi.
RT-PCR usulining o'ziga xosligini gibridizatsiya problarini reaksiya aralashmasiga kiritish orqali oshirish mumkin. Ushbu zondlar lyuminestsent bo'yoqlar yoki lyuminestsent bo'yoqlar va floresan söndürücüler kombinatsiyasi bilan etiketlanishi kerak. Taqman usulida Taq polimerazasining 5'-3' ekzonukleaza faolligi qo'llaniladi, bu primerlar orasidagi kuchaytirilgan DNK hududiga maxsus bog'langan va polimerizatsiya jarayonining normal jarayoniga xalaqit beradigan etiketli oligonukleotid zondini yo'q qiladi. Ushbu yondashuv ikki yorliqli ftorogen gibridizatsiya zondlaridan foydalanadi. Bitta yorliq lyuminestsent bo'yoq bo'lib, muxbir bo'lib xizmat qiladi, ikkinchi yorliq muxbir bo'yog'ining emissiya spektrini so'ndiruvchi bo'yoqdir. Gibridizatsiya zondining nukleaz degradatsiyasi muxbir bo'yog'ini söndürücü bilan bog'lashdan ozod qiladi, natijada uning floresan emissiyasi maksimal darajaga ko'tariladi. Qurilma tomonidan aniqlangan floresansning oshishi ma'lum bir PCR mahsuloti olinganligini bildiradi va floresan intensivligi mahsulot miqdoriga bog'liq.
Shunday qilib, RT-PCR usulining muhim xususiyati, klassik PCRdan farqli o'laroq, sinov materialida yuqumli agentlarning DNK / RNK ni miqdoriy aniqlash imkoniyati, elektroforez bosqichining yo'qligi, davolashni tashkil etish uchun kamroq qat'iy talablardir. PCR laboratoriyasi va olingan natijalarni avtomatik ro'yxatga olish va talqin qilish.
Elektroforez bosqichining yo'qligi tadqiqotga sarflangan vaqtni qisqartirish va atrof-muhitni PCR mahsulotlari bilan ifloslanish (ifloslanish) xavfini minimallashtirish va shu bilan noto'g'ri ijobiy natijalar sonini keskin kamaytirish imkonini beradi.
7. Biotexnologiya va genetik muhandislik.
Asrimizning 60-yillari o'rtalarida "yangi biologiya" paydo bo'ldi, uning amaliy sohalari rivojlanishi bir qator kimyoviy va farmatsevtik vositalarni olish tartiblarini sezilarli darajada o'zgartirdi. Bu keyingi o'n yillikda biokimyo, genetika, hujayra biologiyasi va molekulyar biologiya bo'yicha ko'plab kashfiyotlar tufayli haqiqatga aylandi. Bu kashfiyotlar hujayralardagi DNK ning tuzilishi va funksiyasini aniqlash, ko‘plab fermentlarning tuzilishi va funksiyalarini ochishni o‘z ichiga oladi. Ushbu kashfiyotlar tufayli genetik muhandislik usullari ishlab chiqildi. DNK molekulalari bilan manipulyatsiya qilish mumkin bo'ldi: DNK molekulalarini bir organizmdan ikkinchisiga o'tkazish, ilgari tabiatda mavjud bo'lmagan gibrid molekulalarni ishlab chiqarish. Bularning barchasi virusologiya va bakteriologiya, molekulyar biologiya, enzimologiya va boshqa biologiya fanlarining jadal rivojlanishi tufayli mumkin bo'ldi. Bu barcha fanlar chorrahasida biologiyaning yangi yo'nalishi - biotexnologiya paydo bo'ldi . Biotexnologiya, mohiyatiga ko'ra, bakteriyalar, xamirturushlar, hayvonlar yoki o'simliklarning hujayra madaniyatidan foydalanishdan boshqa narsa emas, ularning metabolizmi va biosintetik imkoniyatlari o'ziga xos moddalarni ishlab chiqarishni ta'minlaydi. 1978 yilda tashkil etilgan Evropa biotexnologiya federatsiyasining ta'rifiga ko'ra, biokimyo, mikrobiologiya, genetika va kimyoviy texnologiya bilimlari va usullarini qo'llashga asoslangan biotexnologiya texnologik jarayonlarda mikroorganizmlar va hujayra madaniyatining xususiyatlaridan foydalanishga imkon beradi. . Bu odamlarning hayoti va farovonligi uchun muhim bo'lgan moddalar va birikmalarni osongina olinadigan va qayta tiklanadigan resurslar yordamida olish imkoniyatini yaratadi.
Sanoat miqyosida bunday biotexnologiya allaqachon biosanoatni tashkil etadi. Biotexnologiya an'anaviy sanoatda va biotexnologiyaning o'zi etakchi rol o'ynaydigan sohalarda qo'llanilishi mumkin. An'anaviy sanoatda biotexnologiya oziq-ovqat sanoatida (sun'iy lazzatlar sintezi), to'qimachilik (ayrim polimerlar va xom ashyo ishlab chiqarish), energiya (metanol, etanol, biogaz va vodorod ishlab chiqarish) va hatto metallurgiyada (ba'zi metallarni olishda) qo'llaniladi. ). Sanoatning ikkinchi guruhida biotexnologiya quyidagilarni o'z ichiga oladi:
1. Oziq-ovqat ishlab chiqarish (oqsillar, aminokislotalar, vitaminlar ishlab chiqarish, shuningdek, ularning fermentlaridan foydalanish uchun xamirturush, suv o'tlari va bakteriyalarni keng ko'lamda etishtirish).
2. Qishloq xoʻjaligi (oʻsimlik navlarini klonlash va tanlash, bioinsektitsidlar ishlab chiqarish, yuqori mahsuldor hayvonlarni klonlash).
3. Farmatsevtika sanoati (vaksinalar, gormonlar, interferonlar va antibiotiklarni ishlab chiqish va sintez qilish).
4. Atrof-muhitni muhofaza qilish va uning ifloslanishini kamaytirish (oqava suvlarni tozalash, maishiy chiqindilarni qayta ishlash, kompost ishlab chiqarish, shuningdek, mikroorganizmlar tomonidan parchalanishi mumkin bo'lgan birikmalar ishlab chiqarish).
Ehtimol, eng qadimgi biotexnologik jarayon mikroorganizmlar yordamida fermentatsiya edi. 1981 yilda Bobil Balada olib borilgan qazishmalar paytida miloddan avvalgi 6-ming yillikka oid loydan yasalgan lavha topilgan. Unda pivo tayyorlash retsepti yozilgan edi. Fermentatsiya jarayonlarini takomillashtirish va ularning samaradorligini oshirish zamburug'lar va mikroorganizmlar hujayralaridan turli xil moddalarni, masalan, antibiotiklar va antiparazitar preparatlarni ajratib olish usullarini ishlab chiqish bilan parallel ravishda amalga oshirildi.
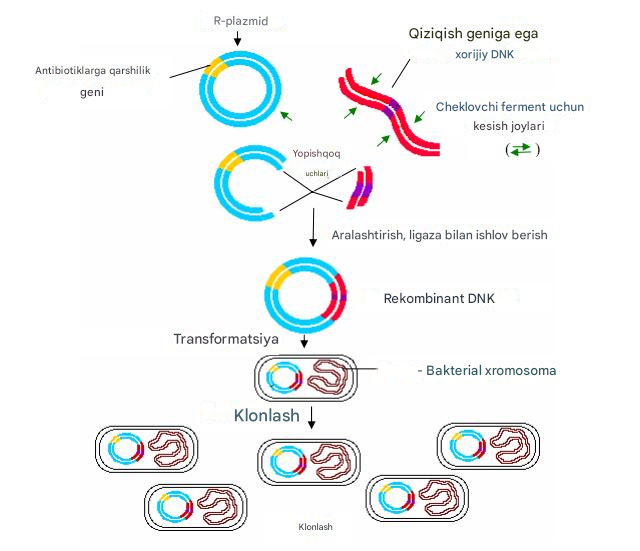
genetik muhandislik usullariga asoslangan . Gen muhandisligining asosini rekombinant DNK molekulalari bilan tajribalar tashkil etadi. AQSh Milliy Sog'liqni saqlash instituti ma'lumotlariga ko'ra, rekombinant DNK - bu tirik hujayradan tashqarida, sinov naychasida, tabiiy yoki sintetik DNK bo'laklarini hujayrada ko'paya oladigan molekulalar bilan birlashtirish orqali hosil bo'lgan DNK molekulalari. Tajribaning asosi - plazmid yoki virus genomi bo'lgan vektorga tabiiy yoki begona DNKning integratsiyasi. Keyin rekombinant DNK molekulasi odatda bakterial hujayra ichiga kiritiladi va u erda replikatsiya qilinadi. Bu jarayon transformatsiya deb ataladi. Bunday rekombinant molekulani o'z ichiga olgan hujayra ko'payib, o'zgartirilgan hujayralar klonini hosil qiladi. Biotexnologiyaning maqsadlaridan biri ko'p miqdorda bakteriyalar uchun begona o'ziga xos oqsillarni ishlab chiqarishga qodir transformatsiyalangan hujayralar klonlarini olishdir.
12-rasm. Rekombinant DNKni tayyorlash va klonlash
Klonlash usuli keng qo'llanilishini topdi. Uning yordami bilan siz ishlatiladigan mikrobiologik mahsulotlarni olishingiz mumkin odam. Hozirgi vaqtda insulin gormonini mikrobiologik ishlab chiqarish usullari ishlab chiqilgan. Ilgari, uni sigir va cho'chqalarning oshqozon osti bezidan ajratib olish yo'li bilan olingan, bu juda qiyin va qimmat. Interferon va somatotropin olish usullari ishlab chiqilgan. Bu moddalarning sintezini belgilovchi genlar asosan E. coli hujayralarida klonlangan. Genetik injeneriya usullaridan foydalangan holda tibbiy va veterinariya maqsadlarida ishlatiladigan yuzlab dori vositalari yaratildi va rekombinant super ishlab chiqaruvchi shtammlar olindi, ularning ko'pchiligi amaliy qo'llanilishini topdi. Tibbiyotda gepatit B, interleykinlar-1, 2, 3, 6, insulin, o'simta nekrozi omili, timik peptidlar, miyelopeptidlar, to'qimalarning plazminogen faollashtiruvchisi, eritropoetin, OIV antijenleri, qon ivish faktorlari, monoklonli antikorlar va ko'plab antikorlarga qarshi genetik ishlab chiqilgan vaktsinalar allaqachon qo'llaniladi. diagnostik maqsadlar uchun antijenler.
Xulosa
Xulosa qilib aytganda, genetik muhandislik istiqbollari haqida bir necha so'z aytish kerak, bu shunchaki ajoyib. Gen injeneriyasining imkoniyatlari va real yutuqlarini batafsil tahlil qilish asosida XXI asr o‘rtalari uchun ilmiy prognozlar tuzildi. Masalan, kelajakda OITS kabi xavfli kasallikni davolash uchun dori vositalari ishlab chiqilishi, xavfli oʻsmalarga aloqador genlar aniqlanishi, saratonning deyarli barcha turlarining paydo boʻlish mexanizmlari yoʻlga qoʻyilishi va rivojlanishiga umid bildirildi. saraton kasalligini oldini oluvchi dori vositalari tugallanadi. Hozirgi vaqtda bir qator mamlakatlarda, shu jumladan Rossiyada inson biologiyasi, evolyutsiyasi, biologiyasi bo'yicha muhim kashfiyotlar va'da qilingan "Inson genomi" xalqaro dasturi ishlab chiqilmoqda.
rivojlanish va neyrobiologiya.
O'z-o'zini tayyorlash uchun vazifalar (o'z-o'zini nazorat qilish)
O'z-o'zini nazorat qilish uchun savollar:
Mikroorganizmlar genetikasi va uning biologiya fanlari tizimidagi o'rni.
Bakteriyalarda genetik materialni tashkil qilish xususiyatlari.
Xromosomadan tashqari genetik elementlar - plazmidlar. Ularning tasnifi va mikroorganizmlar evolyutsiyasidagi roli.
Migratsiya qiluvchi genetik elementlar, ularning tasnifi. Migratsiya elementlari va mikroorganizmlar evolyutsiyasi.
Mikroorganizmlarda fenotip va genotip haqida tushuncha.
Mutatsiyalar. Bakteriyalardagi mutatsiyalarning turlari va mutantlarni aniqlash usullari.
Bakteriyalarda rekombinatsiya. Bakteriyalarda irsiy axborot almashish usullari, ularning umumiy xususiyatlari
Konjugatsiya, jarayonning umumiy xususiyatlari. Donorlarning turlari.
Transformatsiya. Transformatsiya uchun shartlar.
Transduktsiya, uning turlari. O'tkazuvchi faglarning hosil bo'lish mexanizmlari.
Yuqumli kasalliklarni aniqlashning molekulyar genetik usullari. PCR, RT-PCR.
Biotexnologiya. Genetika muhandisligi
Jadvalni to'ldiring:
Bakteriyalarda genetik almashinuv
Genetik materialni uzatish usuli
ta'rifi
Genetik almashinuv ishtirokchilarining xususiyatlari








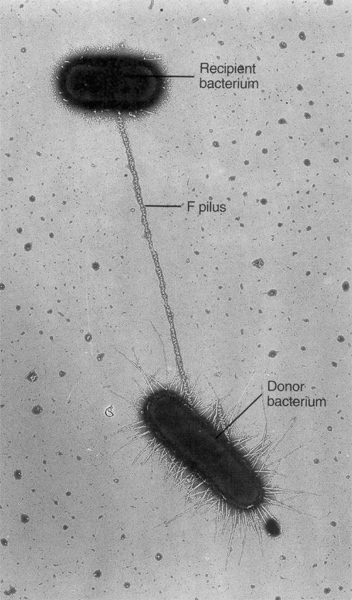 -rasm. Bakterial konjugatsiyaning mikrografisi.
-rasm. Bakterial konjugatsiyaning mikrografisi.